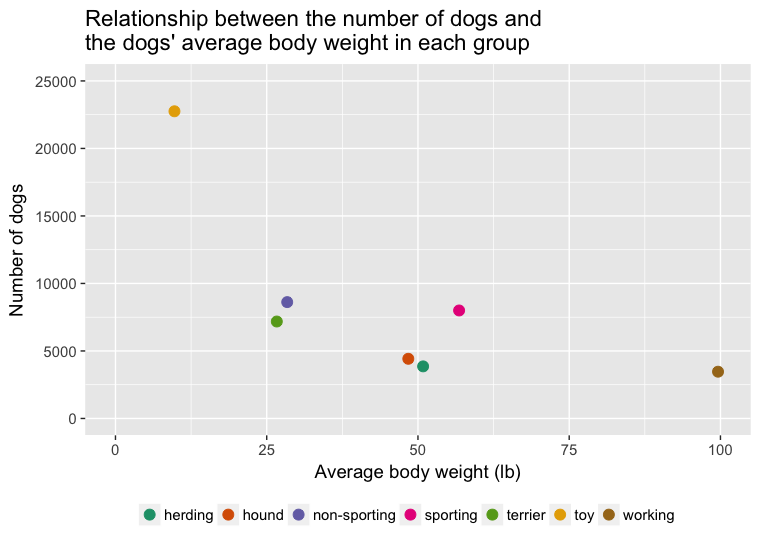
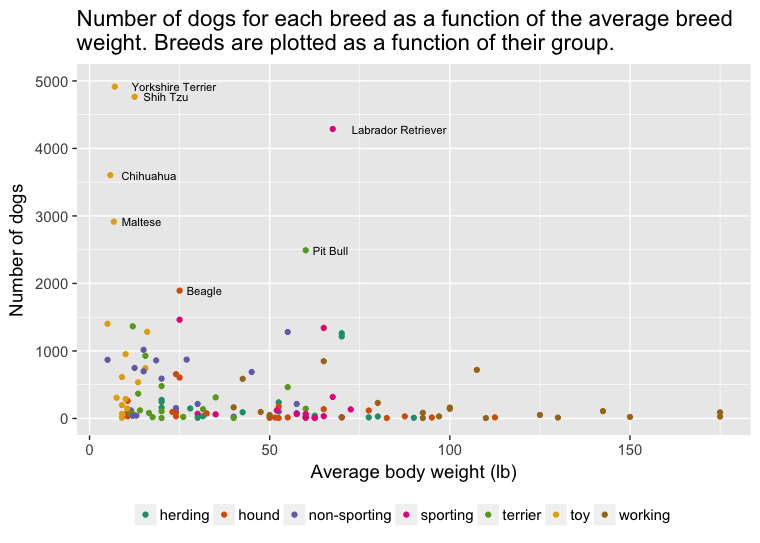
Sara Shettleworth (2010) dans son ouvrage intitulé Cognition, Evolution and Behavior, aborde brièvement la capacité d’un individu à exercer une surveillance ainsi qu’un contrôle sur ses propres capacités cognitives. L’étude de ces capacités chez les êtres humains ainsi que chez les animaux non-humains1 provient du domaine de la métacognition. Bien que les capacités métacognitives des animaux soient étudiées chez le rat (Foote et Crystal, 2007) et le pigeon (Sutton et Shettleworth, 2008), la majorité des données proviennent d’études effectuées chez les primates (Marsh et MacDonald, 2012) et les macaques (Hampton, 2001).
Récemment, McMahon, Macpherson et Roberts (2010) ont élargi l’étendue des espèces en s’interrogeant sur la capacité du chien domestique à utiliser un résonnement métacognitif dans une tâche de recherche d’informations. Dans ce type de tâche, un individu doit retrouver de la nourriture cachée à l’intérieur d’un contenant faisant partie d’un ensemble plusieurs autres contenants. Deux conditions sont habituellement présentées. Dans une première condition, l’individu voit directement la nourriture être cachée à l’intérieur du contenant avant de pouvoir faire son choix alors que dans une seconde condition l’individu n’est pas en mesure de voir où est cachée la nourriture. L’individu a aussi la chance d’émettre une réponse qui lui permet d’obtenir de l’information concernant l’endroit où est cachée la nourriture, et ce, dans les deux conditions. La logique derrière l’utilisation de cette tâche est simple. L’animal va émettre la réponse qui lui permet d’obtenir de l’information concernant l’endroit où la nourriture est cachée dans la condition où il ne voit pas la nourriture être directement déposée à l’intérieur de l’un des contenants. À l’opposé, l’animal va directement chercher la nourriture dans le bon contenant lorsque celui-ci voit la nourriture y être directement déposée, sans émettre le comportement de recherche d’informations. Ce patron de recherche s’expliquerait par la capacité de l’individu à sonder sa mémoire pour y retrouver de l’information par rapport à l’endroit où se cache la nourriture. Lorsque cet exercice de surveillance est infructueux et aucune information n’est disponible en mémoire, comme c’est le cas dans la condition où l’animal n’est pas en mesure de voir la nourriture être cachée, l’individu apporte les correctifs nécessaires et cherche à obtenir plus d’informations. Cette recherche d’informations se traduit par l’animal qui choisit d’émettre une réponse qui lui permet de prendre connaissance d’où ce trouve la nourriture. En se basant sur cette logique, McMahon et ses collaborateurs testent la capacité du chien domestique à utiliser un raisonnement métacognitif.
Six chiens domestiques appartenant à des propriétaires privés prennent part à une première expérience. Avant le début de la phase de test, les chiens sont progressivement entrainés à retrouver de la nourriture à l’intérieur de l’une de deux, trois et finalement quatre boites. Afin de permettre aux chiens de distinguer la boite contenant la nourriture des autres boites, l’un des côtés de la boite (perpendiculaire à l’animal) est toujours blanc. Les autres boites, quant à elles, sont toutes noires (voir Figure 1). La phase de test se distingue des phases d’entrainement par une rotation des boites faisant progressivement passer le côté blanc à un angle de 0°, 45°, 90° et135 ° par rapport à la position de départ du chien. Le côté blanc est toujours visible de la position de départ dans les essais où l’angle est de 0° ou 45°, visible selon la position de la boite dans l’ensemble de quatre boites lorsque l’angle est de 90° et finalement impossible à voir de la position de départ lorsque l’angle est de 135°. Suite aux sessions où l’angle est de 135°, huit essais guidés sont présentés aux chiens.
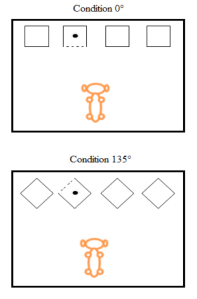
Figure 1. Figure présentant la situation expérimentale aux essais tests de la première condition. Le point noir représente la nourriture et la ligne pointillée le côté de la boite de blanc.
Dans ces essais, un expérimentateur déplace le chien pour que celui-ci soit en mesure d’apercevoir quelle boite présente le côté blanc, le chien est ensuite relâché et doit faire son choix. Bien que les auteurs ne mentionnent pas les raisons de l’introduction des essais guidées, j’ai l’impression que ceux-ci sont instaurés pour écarter la possibilité que les chiens ne comprennent tout simplement pas qu’aux essais où l’angle est de 135°, le côté blanc (qui indique la position de la nourriture) n’est visible que du côté inverse à la position de départ d’où est relâché le chien (voir Figure 1). Faisant suite à ces essais, 16 nouveaux essais avec un angle de 135° sont administrés. Dans ces essais, le chien est maintenant conscient (ou du moins c’est ce que les auteurs croient) qu’une réorientation comme celle introduite lors des essais guidés lui permet de retrouver le côté blanc, un indice crucial pour être en mesure de retrouver la nourriture. Donc l’absence de comportement de recherche d’informations dans ces essais peut maintenant être plus facilement interprétée comme l’incapacité des chiens à utiliser un résonnement métacognitif pour effectuer un comportement de recherche d’informations qui lui permettrait de retrouver la nourriture, ce qui n’était pas le cas aux essais à 135° avant l’introduction des essais guidés.
Les chiens performent au-dessus du niveau de la chance pour les essais à 0° et 45° indiquant que ceux-ci sont en mesure de comprendre la tâche lorsqu’ils voient le côté blanc de la boîte. Cependant, les chiens performent au niveau de la chance dans les essais à 90° et 135°. La performance des chiens se retrouve au dessus du niveau de la chance dans les 8 essais guidés pour de nouveau retomber au niveau de la chance dans les 16 essais repris à 135°. Ces résultats démontrent donc que dans les conditions où le côté blanc n’est pas visible (90° et 135°), les chiens domestiques ne cherchent pas à obtenir de l’information sur l’emplacement de la boite présentant le côté blanc, et ce, même après des essais réussis où ils sont réorientés par un expérimentateur. Comme le notent les auteurs, il est possible que les chiens domestiques aient des capacités métacognitives, mais qu’ils aient tout simplement choisi de les ignorer au profit d’un patron de performance reflétant un taux de réponse moindre, mais rapide. En effet, les auteurs argumentent qu’il est possible que les chiens choisissent de ne pas se réorienter puisque ce comportement introduit un délai dans l’obtention de la récompense. Les auteurs choisissent donc d’introduire une nouvelle tâche qui cette fois-ci contrôle pour l’introduction d’un délai.
Dans leur deuxième expérience, les auteurs administrent à 7 chiens une tâche en deux phases. Dans une première phase, le chien doit choisir entre deux expérimentateurs positionnés devant lui, un expérimentateur qui fournit une information sur la boite contenant de la nourriture dans un ensemble de trois boites placées parallèlement à la position de départ du chien et un deuxième expérimentateur qui ne donne aucun indice. Dans la deuxième phase, le chien voit l’expérimentateur qui donne l’information pointer la boite dans laquelle la nourriture est cachée si celui-ci a été choisi par le chien lors de la première phase. Lorsque le chien choisit l’expérimentateur qui ne fournit pas d’information, il voit tout simplement l’expérimentateur lui faire dos pour une durée équivalente au temps où l’autre expérimentateur pointe la boite contenant la nourriture. Le rôle de chaque expérimentateur est contrebalancé ainsi que le côté où ils se trouvent pour chacun des essais.
Dans la première phase de cette tâche, 6 des 7 chiens ont choisi au-delà du niveau de la chance l’expérimentateur qui fournissait de l’information par rapport à la boite où la nourriture se cachait. Dans la seconde phase, la proportion de réponses correctes (choisir la boite contenant de la nourriture) lorsque les chiens choisissent l’expérimentateur qui fournit de l’information est supérieure à ce qui est attendu par le hasard alors qu’elle ne diffère pas du niveau de la chance lorsque les chiens choisissent l’expérimentateur qui leur fait dos. Ces résultats semblent être concordants avec l’utilisation de capacités métacognitives de la part des chiens puisque ceux-ci, n’étant pas au courant de l’endroit où est cachée la nourriture, préfèrent l’expérimentateur qui leur fournit un indice leur permettant d’identifier le contenant où la nourriture se cache à celui qui ne leur fournit pas d’information concernant la position de la nourriture. Cependant comme les auteurs l’argumentent, il est possible que les chiens préfèrent tout simplement l’expérimentateur qui fournit de l’information puisqu’il est associé à une plus grande probabilité d’obtenir une récompense (72,6% versus 36,3% pour l’expérimentateur qui leur fait dos).
Pour contrôler pour cette possibilité, les auteurs administrent à sept nouveaux chiens domestiques une troisième expérience qui diffère sur un point important : peu importe le choix d’expérimentateur la probabilité d’obtenir un renforcement est maintenant identique. Pour ce faire, lorsque le chien choisit l’expérimentateur qui leur fait dos, de la nourriture est cachée sous tous les contenants. Cela fait donc en sorte que le chien est certain de trouver de la nourriture, peu importe, son choix de contenant. Étrangement, lorsque l’animal choisit l’expérimentateur qui pointe la boite contenant la nourriture, de la nourriture est seulement cachée sous la boite qui sera ensuite pointée par l’expérimentateur. Les auteurs justifient cette disparité en mentionnant que les chiens ont très bien performé à cette condition (94,5%). Cependant, il était impossible de prédire la performance des chiens avant le début de l’Expérience 3. Par conséquent, ce choix de procédure de la part des auteurs demeure étrange étant donné que la performance lors du choix de l’expérimentateur qui fournit de l’information à l’Expérience 2 n’était que de 72,6%.
Dans cette troisième expérience, tous les chiens ont préféré choisir l’expérimentateur qui fournit de l’information concernant la boite contenant de la nourriture. Tout comme lors de la deuxième expérience, la proportion de réponses correctes lorsque les chiens choisissent l’expérimentateur qui fournit de l’information est supérieure à ce qui est attendu par le hasard et la proportion de réponses correctes ne diffère pas du niveau de la chance lorsque les chiens choisissent l’expérimentateur qui leur fait dos. Les résultats de la troisième expérience démontrent clairement que les chiens domestiques sont en mesure, dans une tâche comme celle présentée par les auteurs, de choisir au-delà du niveau prédit par la chance, l’expérimentateur qui lui fournit de l’information concernant l’endroit où se cache de la nourriture dans un ensemble de trois boites.
Maintenant que ce fait est établi, il est important de se demander si ce patron de résultat est une preuve de l’utilisation de capacité métacognitive chez le chien domestique. Je crois personnellement que cette procédure, malgré l’ajout d’une troisième expérience, est loin d’être une démonstration claire d’une telle capacité chez le chien domestique. Mon argumentation est basée sur le fait qu’il subsiste un important biais dans la procédure utilisée par les auteurs qui fait en sorte que le patron de résultat des deux dernières expériences peut être attribuable à un mécanisme d’apprentissage de bas niveau plutôt qu’à l’explication métacognitive de plus haut niveau. En effet, il est possible que la préférence des chiens domestiques pour l’un des deux expérimentateurs soit due aux différences lors de la deuxième phase de la tâche. Il est possible que les chiens préfèrent les situations où un expérimentateur lui pointe la bonne réponse. Le simple fait d’interagir avec un être humain peut être perçu comme plus stimulant de la part du chien ce qui en retour augmente l’attrait de cette situation. Tout au contraire, la situation où l’expérimentateur fait dos au chien est beaucoup moins stimulante pour le chien ce qui peut réduire la probabilité que cet expérimentateur soit choisi. En d’autres termes, à probabilité égale d’obtenir un renforçateur (ce qui est argumenté par les chercheurs pour la troisième expérience), les chiens choisissent la situation la plus attrayante sans s’interroger sur le fait qu’ils ne connaissent pas l’endroit où la nourriture est cachée.
En guise de conclusion, il n’est pas impossible que les chiens démontrent des capacités métacognitives, mais pour l’instant, l’étude de McMahon et ses collaborateurs (2010) ne supporte pas de façon claire et nette cette conclusion et d’autres études chez les canidés sont nécessaires pour établir si oui ou non ceux-ci possèdent des capacités métacognitives.
1 Il est à noter que le terme « animal » est utilisé passé ce point pour faire référence à tous les animaux non humains.
Pour ceux qui seraient intéressés à consulter les articles auxquelles je fais référence dans mon commentaire :
Marsh, H. L., & MacDonald, S. E. (2012). Orangutans (Pongo abelii) “play the odds”: information-seeking strategies in relation to cost, risk, and benefit. Journal of Comparative Psychology (Washington, D.C. : 1983), 126(3), 263–78. doi:10.1037/a0025906
Hampton, R. R. (2001). Rhesus monkeys know when they remember. Proceedings of the National Academy of Sciences of the United States of America, 98(9), 5359–62. doi:10.1073/pnas.071600998
Foote, A. L., & Crystal, J. D. (2007). Metacognition in the rat. Current Biology : CB, 17(6), 551–5. doi:10.1016/j.cub.2007.01.061
Sutton, J. E., & Shettleworth, S. J. (2008). Memory without awareness: pigeons do not show metamemory in delayed matching to sample. Journal of Experimental Psychology. Animal Behavior Processes, 34(2), 266–82. doi:10.1037/0097-7403.34.2.266
McMahon, S., Macpherson, K., & Roberts, W. a. (2010). Dogs choose a human informant: metacognition in canines. Behavioural Processes, 85(3), 293–8. doi:10.1016/j.beproc.2010.07.014
Commentaire préparé par Pierre Nadeau-Marchand, étudiant à la maîtrise en psychologie à l’Université de Moncton